Cuprins
- Introducere
- Scurt istoric
- Activitatea catalitică a enzimelor
- Temperatura optimă a reacţiilor enzimatice
- Influenta pH-ului
- Specificitatea enzimelor
- Clasificarea enzimelor
- Lipoxigenaza
- Funcţiile biologice şi clasificarea lipoxigenazelor
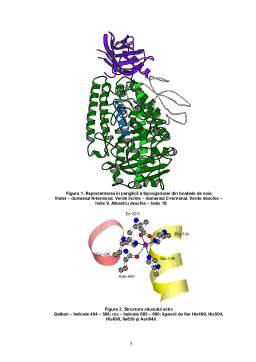
- Structura tridimensională a lipoxigenazei
- Situsul activ
- Reacţiile care au loc la oxidarea acizilor graşi polinesaturaţi
- Modificări de culoare
- Modificări de aromă
- Modificări de textură
- Modificări nutriţionale
- Funcţiuni în vivo ale lipoxigenazei
- Produşii lipoxigenazei
- Activitatea lipoxigenazei în seminţele plantelor
- Metabolizarea acidului arahidonic cu ajutorul lipoxigenazei
- Bibliografie
Extras din proiect
Introducere
În organismele vii se petrec cu o uimitoare uşurinţă, la temperatura joasă şi în soluţie practic neutră, un număr mare de reacţii pe care chimistul nu le poate efectua în laborator decât lucrând la temperaturi şi presiuni ridicate, în prezenţă de acizi sau de baze tari, de dizolvanţi neapoşi sau de catalizatori heterogeni metalici. Printre aceste reacţii se numără atât degradări de molecule (hidrolize şi oxidări), cât şi sinteza de compuşi cu structură complicată. Înţelegerea mersului acestor reacţii este importantă, în primul rând pentru cunoaşterea unor fenomene naturale de cea mai mare amploare şi răspândire, în al doilea rând pentru interesul practic pe care îl prezintă. Nu este absurdă speranţa că, o dată cunoscut mersul reacţiilor din celulele vii, acestea vor putea fi imitate în laborator şi în industrie sau chiar dirijate pe căi noi.
S-a recunoscut încă de mult că organismele folosesc, pentru realizarea acestor transformări chimice, catalizatori organici, conţinuţi în concentraţii mici în celule sau în sucurile secretate de acestea, cum sunt sucurile digestive, laptele, urina etc. S-a dat acestor catalizatori numele de fermenţi sau enzime (de la enzyme, literal: „în aluat”).
Scurt istoric
Reacţiile enzimatice au fost folosite din timpurile cele mai vechi pentru fabricarea vinului, a oţetului, a berii şi a brânzei. O cercetare sistematică a lor a fost întreprinsă abia în epoca modernă.
În 1713, Reamur a observat dizolvarea cărnii în sucul stomacal al ciorii. De asemenea, fiziologul Spallanzani (1783) a hrănit animale cu bucăţi de carne învelite în reţele de sârmă şi observat dizolvarea cărnii în stomac.
Stahl, fondatorul teoriei flagisticului, explică fermentaţia ca un proces în care una din substanţele prezente transmite „mişcarea sa internă” substanţei care fermentează (1697). În 1680, van Leeuwenhoeck a observat la microscop celulele drojdiei de bere, dar această descoperire nu a fost luată în seamă timp de două secole. Lavoisier (1789) a făcut un bilanţ de materiale al fermentaţiei, arătând ca oxigenul, hidrogenul şi carbonul din zahăr se regăsesc în alcoolul şi bioxidul de carbon ce iau naştere.
În cursul sec. al XIX-lea au fost preparate multe extracte de enzime. Astfel, după ce Kirchoff a observat, în 1820, că o componentă glutinoasă din bobul de orz încolţit, numit malţ, transformă cantităţi de amidon mult mai mari decât propria sa greutate, într-un zahăr solubil, maltoza, Dubrunfaut a găsit, în 1830, ca extractul apos, limpede, de malţ are aceeaşi acţiune solubilizantă asupra amidonului ca malţul însuşi. Din acest extract, Payen şi Persoz (1833) au izolat, prin precipitarea cu etanol, prima enzimă, amilaza (fireşte foarte impură), sub forma unui material solid alb, amorf, capabil să solubilizeze o cantitate de amidon de 2000 de ori mai mare decât propria sa greutate. În 1830, Robiquet şi Boutron-Chalard au descoperit hidroliza amigdalinei, cu extract de migdale amare, iar în 1837, Liebig şi Wohler au izolat enzima respectivă, numind-o emulsină. Printre primele enzime izolate (în stare impură) vom mai menţiona: pepsina din sucul gastric (Schwann, 1836); tripsina, din sucul pancreatic (Kuhne, 1848); lipaza (Claude Bernard, 1849); invertaza (Mitscherlich, 1841; Berthelot, 1860); ureaza (Musculus, 1882) etc.
Un moment istoric deosebit de important este recunoaşterea clară, de către Berzelius, în 1835, a caracterului catalitic al reacţiilor enzimatice, precum şi a rolului esenţial pentru viaţa animalelor şi a plantelor jucat de aceste reacţii.
În anul 1940, cercetătorul american Edward Howell a făcut, în acelaşi domeniu, o şi mai mare descoperire: cercetând substanţele vitale propriu-zise, şi anume, enzimele, a dovedit că ele sunt purtătorii vieţii din orice organism viu, fiind deci şi materia vie din alimentele noastre (asta atâta timp cât nu sunt distruse prin fierbere).
Activitatea catalitică a enzimelor
Enzimele sunt, precum s-a mai spus, catalizatori organici, produşi de celula vie, acţionând asupra anumitor substanţe numite substraturi. În marea lor majoritate, enzimele catalizează reacţia unei substanţe organice cu un compus anorganic liber sau cedat de alt compus organic (apă, acid fosforic, hidrogen, oxigen etc.).
Legile catalizei se aplică fireşte şi la enzime. Enzimele, ca toţi catalizatorii, nu catalizează decât reacţii termodinamic posibile, decurgând în sensul stabilirii unui echilibru.
Reacţiile enzimatice prezintă însă unele deosebiri caracteristice faţă de reacţiile catalitice obişnuite, omogene sau heterogene.
Când o reacţie poate fi catalizată atât de o enzimă cât şi de substanţe simple (acizi, baze sau ioni metalici) se constată de obicei că reacţia enzimatică decurge cu viteza mult mai mare; cu alte cuvinte, reacţia enzimatică are o energie de activare mult mai mică. Astfel s-a stabilit că este necesară o concentraţie de ioni de hidrogen de zece milioane de ori mai mare decât de invertază pentru a hidroliza o anumită cantitate de zaharoză, într-un timp dat, la 370.
Temperatura optimă a reacţiilor enzimatice
Viteza reacţiilor enzimatice creşte, ca a celor mai multe reacţii între molecule covalente, cu temperatura, potrivit cunoscutei reguli a lui van Hoff, şi anume, o urcare a temperaturii cu 100 produce o creştere a vitezei de reacţie cu un coeficient 1,5-3. Creşterea acesta se observă însă numai la temperaturi relativ joase. O dată depăşită o anumită temperatură optimă, la care viteza este maximă, aceasta scade, iar la temperaturi mai înalte reacţia încetează. Fenomenul se explică prin faptul, semnalat mai sus, că la temperaturi mai înalte enzimele sunt inactivate prin denaturarea componentei proteice. Cele mai multe enzime devin complet inactive între 50-800. Temperatura optimă nu poate fi însă exact definită, căci ea variază în limite largi, cu concentraţia enzimei, cu concentraţia ionilor de hidrogen şi cu prezenţa diferitelor impurităţi ale preparatului enzimatic sau ale substratului.
Influenta pH-ului
După cum a arătat Sorensen (1909), activitatea enzimelor depinde într-o foarte mare măsura de concentraţia ionilor de hidrogen din soluţie (sau mai corect de activitatea termodinamică a ionilor de hidrogen, adică de pH-ul soluţiei). Curbele reprezentând variaţia vitezei de reacţie cu pH-ul prezintă de obicei un maxim pronunţat la un anumit pH, în timp ce la valori ale pH-ului diferind cu 1 faţă de acest maxim, viteza de reacţie prezintă valori considerabil mai mici. Din cauza acestei particularităţi, este necesar ca în cursul reacţiilor enzimatice să se menţină pH-ul optim constant, prin folosirea unui tampon.
Specificitatea enzimelor
O anumită enzimă catalizează numai un număr mic de reacţii şi de multe ori o singură reacţie, spre deosebire de catalizatori obişnuiţi anorganici (acizi, baze, catalizatori de hidrogenare etc.) care activează practic toate reacţiile posibile de un anumit tip.
Preview document















Conținut arhivă zip
- pagina titlu.doc
- Lipoxigenaza.doc
- cuprins.doc
- referinte.doc
Alții au mai descărcat și

Berea este o bautura alcoolica nedistilata, obisnuita prin fermentarea cu drojdie a unui must realizat din malt, apa si fiert cu hamei. Unii...

Proprietati mecanice - Elasticitatea - Plasticitatea - Duritatea - Rezistenta Proprietati fizice - Electrice - Magnetice - Optice -...
Te-ar putea interesa și

INTRODUCERE Omul primitiv lua cele necesare din mediul înconjurător şi căuta să vindece toate bolile cu ajutorul plantelor , mineralelor şi...
I. ENZIME I.1 SCURT ISTORIC Reactiile enzimatice au fost folosite din timpurile cele mai vechi pentru fabricarea vinului, a otetului, a berii si...

INTRODUCERE Enzimele au fost recunoscute de mult timp ca un mijloc de îmbunătăţire a procesului de procesare sau a proprietăţilor produselor...

GENERALITǍŢI Enzimele au fost recunoscute de mult timp ca un mijloc de îmbunǎtǎţire a procesului de procesare sau a proprietǎţilor produselor...

1.Introducere Enzimele au fost recunoscute de mult timp ca un mijloc de imbunatăţire a procesului de procesare sau a proprietăţilor produselor...

ANTIOXIDANTII prin definitie, sunt aditivi alimentari care prelungesc durata de pastrare a produselor alimentare, prin protejarea lor fata de...

1. Definirea antivitaminelor Conform „Lexicon – Îndrumar pentru Industria Alimentară” vol. I, antivitaminele sunt: „compuşi analogi structural ai...

ANTIOXIDANTII prin definitie, sunt aditivi alimentari care prelungesc durata de pastrare a produselor alimentare, prin protejarea lor fata de...